


La storia genetica è l'insieme delle scoperte effettuate tramite la genetica delle popolazioni, una branca della genetica che analizza la costituzione genetica delle popolazioni mendeliane in termini qualitativi (varianti alleliche presenti all'interno di una popolazione) e quantitativi (frequenze alleliche e genotipiche). Tali scoperte hanno permesso, mediante l'analisi delle parentele e delle differenze geniche, a livello sia delle etnie che delle popolazioni umane, di ricostruire i flussi migratori, gli incroci, l'emersione o l'eliminazione dei caratteri che contraddistinguono le attuali etnie e popolazioni umane, sia nello spazio sia nel tempo, aiutando a ricostruire la storia dell'uomo sin dalla sua comparsa.
Cenni su comparsa dell'uomo moderno e seconda fuoriuscita dal continente africano
Circa sette milioni di anni fa l'assestamento della crosta terrestre produsse l'innalzamento della Rift Valley che attraversa gli attuali stati di Etiopia, Kenya e Tanzania. I venti carichi di piogge provenienti da ovest furono intrappolati dal sollevamento tettonico di una barriera naturale, il Rift africano, provocando un processo di inaridimento verificatosi sull'altopiano omonimo. Il processo di inaridimento portò infatti alla progressiva formazione dell'attuale savana africana in sostituzione della foresta e la popolazione delle protoscimmie africane si trovò ad essere geograficamente separata in due sottopopolazioni dal Rift: il versante ovest rimase lussureggiante e le protoscimmie stanziate su questo lato si adattarono ad un ambiente boscoso, differenziandosi dalle altre e precorrendo le moderne scimmie antropomorfe, mentre le protoscimmie intrappolate sull'altopiano, a causa del lento inaridimento, si adattarono a condizioni ambientali differenti, quali la scomparsa della foresta sostituita dalla savana africana. Tale tipo di ambiente permise l'evoluzione e l'affermazione del genere Homo[1].
Out-of-Africa
Il processo Out-of-Africa (letteralmente: fuoriuscita dall'Africa) è il primo processo migratorio conosciuto, anche se a differenza dei moderni flussi migratori esso fu dovuto al successo dei primi ominidi, i quali poterono espandersi in un ambiente privo di competitori e particolarmente vantaggioso. L'appartenente al genere Homo progenitore della nostra specie, denominato Homo ergaster, si origina in Africa. La sua evoluzione dà origine all'Homo erectus, e colonizzando ad ondate successive l'Eurasia si adatta alle diverse condizioni ambientali, differenziandosi nelle specie Homo heidelbergensis e successivamente in Homo neanderthalensis con caratteristiche carnivore.
Out-of-Africa 2
Alcuni ominidi appartenenti al genere Homo, anziché partecipare al processo Out-of-Africa si stanziarono nella Rift Valley. Da tali ominidi si originò l'Homo sapiens, una cui sottospecie, definita Homo sapiens sapiens (circa 200 000 anni fa), migrò per affermarsi successivamente come unica specie appartenente al genere Homo[2], e sostituendosi, per motivi tuttora ampiamente dibattuti, sia alle specie comparse a seguito del processo Out-of-Africa, sia a quelle coevolutesi in Africa.
Cronologia del popolamento (Y-DNA)

In genetica umana, il cromosoma Y viene suddiviso in aplogruppi definiti sulla base della mutazione di un singolo nucleotide[3] nella sequenza non ricombinante del cromosoma Y chiamata NRY.[4] Ogni mutazione corrisponde ad un aplotipio e il cromosoma Y viene ereditato di padre in figlio. Dal momento che la mutazione colpisce una sequenza non ricombinante (cioè che non subisce modificazioni quando viene ereditata), è possibile risalire, andando a ritroso di generazione in generazione, alla linea di discendenza maschile.

Aplotipi europei legati all'Y-DNA
Lo studio degli aplotipi ovvero della combinazione delle varianti alleliche lungo un cromosoma o lungo un segmento cromosomico contenente loci strettamente associati tra di loro, e che in genere, vengono ereditati insieme, ha permesso di identificare due aplotipi europei definiti Eu18 ed Eu19 i quali hanno permesso, tramite metodiche di comparazione delle sequenze, di identificare tracce di migrazioni di popolazioni Europee risalenti all'epoca paleolitica, si ritiene che le migrazioni delle popolazioni europee siano dovute a fenomeni ambientali, quali glaciazioni, competizione fra popolazioni e ricerca di cibo, essendo gli aplotipi ereditati insieme, sono stati identificati due nuclei isolati di popolazioni, rispettivamente i nuclei nella penisola Iberica e i nuclei in Ucraina, questi aplotipi costituiscono il 50% dei cromosomi Y europei. L'aplotipo Eu19 è diffuso anche nel Pakistan settentrionale e nell'Asia centrale a supporto dell'ipotesi che queste due popolazioni siano migrate sia verso il centro Europa che verso l'Asia[5].
Aplotipi italiani legati all'Y-DNA
Distribuzione percentuale degli aplotipi del cromosoma Y italiani[6].
| Regione | I1 | I2a | I2b | R1a | R1b | G2a | J2 | J1 | E1b1b | T + (L) | Q |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Italia settentrionale | 6% | 2.5% | 2.5% | 3.5% | 55% | 2.5% | 11.5% | 0.5% | 11% | 4.5% | 0% |
| Italia centrale | 3% | 2% | 5% | 3.5% | 43% | 8.5% | 19.5% | 2% | 10% | 3.5% | 0% |
| Italia meridionale | 2.5% | 2.5% | 2.5% | 2.5% | 29% | 8.5% | 23.5% | 5% | 18% | 5.5% | 0% |
| Sicilia | 3% | 1% | 1% | 4.5% | 30% | 5.5% | 26.5% | 4% | 17.5% | 6% | 1% |
| Sardegna | 0% | 37% | 0% | 0% | 22% | 15% | 10% | 2.5% | 10% | 1.5% | 2% |
Cronologia del popolamento (mtDNA)
Similarmente al cromosoma Y il DNA mitocondriale (mtDNA) contenuto nei mitocondri, viene ereditato da parte materna, consente quindi di risalire alla via patrilineare femminile, l'aplogruppo mitocondriale U5b3 ha permesso di identificare un effetto del fondatore verificatosi circa 10.000 anni fa in Italia, si ritiene che le femmine portatrici siano successivamente migrate in Provenza, probabilmente fra i 9000 e 7000 anni fa, dove si sviluppò la variante U5b3a1. Fenomeni di migrazione successivi avrebbero poi permesso l'introduzione dell'aplotipo U5b3a1 dalla Provenza alla Sardegna, presumibilmente seguendo i commerci di ossidiana, ad oggi circa il 4% della popolazione femminile in Sardegna appartiene a questo aplotipo.[7]
Aplotipi legati all'mtDNA
L'aplogruppo più comune in Europa e in Italia risulta essere l'aplogruppo H originatosi probabilmente circa 20.000 anni fa in Europa meridionale e nel Vicino Oriente, sempre in Europa circa 15.000 anni fa in Spagna si differenzia l'aplogruppo V. L'aplogruppo J si ritiene essersi originato nel Vicino Oriente o nel Caucaso mentre nel nord-est circa 25.000 anni fa si origina l'aplogruppo W, l'aplogruppo T si origina in Mesopotamia circa 17,000 anni fa, l'aplogruppo U si origina in Asia occidentale circa 60,000 anni fa, l'aplogruppo I circa 30.000 anni fa probabilmente in Europa, l'aplogruppo K circa 16.000 anni fa nel Vicino Oriente, l'aplogruppo X2 oltre 30.000 anni fa nel nord-est europeo[8][9]
Tabella percentuale degli aplogruppi femminili di mtDNA presenti in Italia:
| H | V | J | T | U | K | I | W | X2 | altri | |
|---|---|---|---|---|---|---|---|---|---|---|
| % | 33,5% | 4,5% | 7,5% | 12% | 12,5% | 7% | 2% | 2,5% | 2% | 16,5% |
Migrazioni umane riscontrate tramite analisi dell'Y-DNA
Paleolitico superiore
Attraverso gli studi sulla variazione genetica del cromosoma Y in Europa, è stato riscontrato un primo periodo di colonizzazione in epoca tardo-glaciale, la quale ha portato alla differenziazione dei primi aplotipi europei associati all'uomo moderno, ogni aplotipo viene di norma associato ad una popolazione la quale, stanziatasi sul territorio, ha avuto un tempo sufficiente per differenziarsi dalla popolazione di origine, dando via ad un proprio aplotipo caratteristico, l'aplotipo I1a risulta maggiormente presente in Scandinavia, la sua frequenza diminuisce rapidamente spostandosi verso la pianura orientale e la frangia atlantica, tuttavia le differenze riscontrate a livello dei microsatelliti mostrano che probabilmente nel sud della Francia si debba ricercare la zona di origine delle popolazioni portatrici degli aplotipi I1a e I1c. l'aplotipo I1b si estende dall'adriatico orientale fino all'Europa orientale, e diminuisce in direzione dei Balcani meridionali, scompare bruscamente nel nord Italia. Si suppone che l'aplotipo I e sotto prodotti si sia originato durante l'ultima glaciazione nell'area dei Balcani o dell'Europa dell'est. Al contrario, la popolazione portatrice di I1b2, molto probabilmente si originò nel sud della Francia, subendo un'espansione nel periodo post-glaciale, ed a seguito dei flussi migratori, avrebbe poi colonizzato la Sardegna circa 9000 anni fa[10].
Mesolitico e Neolitico

Durante questo periodo si ha un grado di espansione apprezzabile dell'uomo moderno nella penisola Italiana, gli studi attuali risultano molto complessi a causa della deriva genetica e degli effetti fondatori locali che si sono verificati su tutta la penisola in questo periodo di tempo. L'Aplogruppo R1b (Y-DNA), viene ritenuto essere la più antica linea genetica presente nel continente europeo[5], la sua presenza viene comunemente associata ad un effetto del fondatore verificatosi nell'Europa centro occidentale[12][13]. Le popolazioni stanziatesi in Italia dal Mesolitico sono caratterizzate da alte frequenze di R1*(xR1a1), condizione che si ritrova ad oggi nelle popolazioni basche, ritenute le più somiglianti geneticamente ai primi europei, durante il Neolitico i migranti introducono le varianti E3B1 e J2, il 27% delle variazioni genetiche totali, basate sull'analisi dei polimorfismi indicano un chiaro gradiente di distribuzione della popolazione italiana sull'asse nord-sud della penisola, le variazioni introdotte nel Neolitico non sembrano essere dovute a flussi migratori provenienti dalla Spagna, ma si configurano come migrazioni provenienti dall'Asia o dall'Anatolia attraverso l'attuale area Balcanica; diversi autori hanno suggerito che l'asse di distribuzione Nord-Sud delle differenze genetiche fra le popolazioni italiane siano dovute agli eventi di colonizzazione greca nel Sud, tuttavia nuovi studi suggeriscono che in epoca Neolitica fu l'influenza delle popolazioni provenienti dall'Anatolia la causa principale delle differenze nel bacino genetico italiano, assegnando ai greci un ruolo di secondaria importanza; attualmente si assume che durante il Neolitico si consolidò l'aplotipo principale R1*(xR1a1) mentre gli aplotipi HGS, E3B1 e J2 risultano assenti o presenti a bassa frequenza, in particolare nel nord d'Italia si ritrova a bassissima frequenza E3b2 di origine africana[14].
R1b-S28 (U152)
La distribuzione della variazione STR di R1b in Europa diminuisce da est a ovest, suggerendo un ingresso in Europa dall'Asia occidentale con la diffusione dell'agricoltura[15]. R1b viene spesso citato come esempio di una distribuzione ad onda dell'aplogruppo, in questo caso, da est a ovest.[16]. La proposta di un'origine a sud-est di R1b origina con la scoperta dei primi subcladi di R1b localizzati in Asia occidentale e il più recente in Europa occidentale[17][18]. Tuttavia non vi è accordo sulle datazioni della migrazione o migrazioni responsabili di questa distribuzione, non si escludono migrazioni prima o dopo il Neolitico[17].
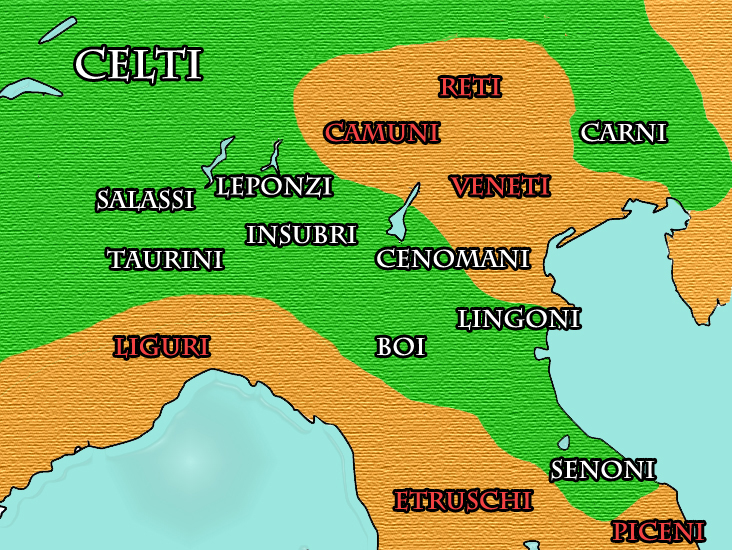
David K. Faux ha ipotizzato che la mutazione S28 (marcatore U152), e relativi sottogruppi, possano essere associati in maniera non esclusiva alla popolazione celtica dell'arco Alpino[19][20].
Alcuni studi hanno identificato l'esistenza di un sottoclade a livello dell'arco Alpino localizzato nell'Italia nord occidentale chiamato S28/U152 il quale suggerisce l'esistenza di un nucleo di dispersione aplotipica proprio della penisola italiana a livello delle Alpi, comprendente il nord-ovest italiano a partire dal 1200 aC[21], tuttavia non è ancora stata identificata la prima area geografica nella quale sia comparso tale marcatore.
Questa subclade dell'aplogruppo R1b è la più comune nella penisola italiana (26,6% al Nord e 10,5% al Sud) e nelle isole di Sardegna e Corsica.
E1b1b (ex E3B)
E1b1b (E-M215), precedentemente noto come E3B (o "aplotipo V")[22] è definito dalla mutazione M215.[23][24][25], attualmente è oggetto di numerose controversie.
Attualmente le linee partilineari di E-M215 ed E-M35 appaiono identiche, si ritiene che l'aplogruppo compaia in Africa orientale circa 22400 anni fa.[23][26]
In Europa è presente E-M35, alcuni studiosi hanno ipotizzato che questa mutazione possa rappresentare il marcatore di un'antica migrazione avvenuta nel tardo Pleistocene dal Nord Africa verso l'Europa, Sinai ed Egitto[27].
In Italia prevale il sottoclade E1b1b1b1 (E-M81).
Alcuni studiosi hanno avanzato l'ipotesi secondo la quale l'aplogruppo E3b1a2 sia stato introdotto in Gran Bretagna a partire dalle popolazioni Caucasiche, spinte in quanto arruolate dall'esercito Romano a colonizzare l'isola Britannica, assegnando alla dominazione imperiale Romana un ruolo indiretto nella diffusione di questo aplotipo, appartenente alla penisola caucasica, nell'isola Britannica[28].
J2a-M410 e J2b-M12
La diffusione dell'aplotipo J2 nel bacino del Mediterraneo viene spesso associata all'espansione dei popoli agricoli durante il periodo Neolitico[29]. La comparsa di J2 è stimata a circa 18,500 anni fa con uno scarto di 3.500 anni fa[30]. Taluni studiosi lo inseriscono fra gli aplogruppi dell'Asia occidentale e sud-orientale, associandolo alla presenza di reperti archeologici del neolitico, come statuette e ceramiche dipinte[31] è stata avanzata l'ipotesi che il subclade J2a-M410 appartenga ai primi agricoltori[32]. Tuttavia altri studiosi ipotizzano un possibile evento di dispersione nel post-neolitico, in particolare legato alla dominazione della Grecia antica[33]. In Europa, la frequenza di aplogruppo J2 scende drammaticamente muovendosi verso nord dal Mediterraneo.
In Italia, J2 si presenta con frequenze regionali che variano tra il 9% e il 36%[34].
È stato proposto che il subclade J2a-M410 sia collegato alle popolazioni dell'antica Creta. L'aplogruppo J2b-M12 è stato associato con la Grecia del Neolitico (circa 8500 - 4300 aC) ed è stato segnalato all'interno di siti Cretesi (3,1%)[35].
J1 J-M267
Negli studi meno recenti viene denominato UE10, questo aplogruppo si trova con frequenze importanti nel Medio Oriente, Caucaso, Nord Africa, Corno d'Africa. Si trova anche meno frequentemente, ma ancora occasionalmente in quantità significative, in Europa e in Estremo Oriente come il subcontinente indiano e nell'Asia centrale.
J1 viene diviso in diversi sub-cladi, alcuni dei quali sono stati riconosciuti prima ancora di J1, per esempio J-M62[36]. Con la sola eccezione di J1c3, la maggior parte dei subcladi non risultano comuni[37]. La frequenza e la diversità di J1 (e anche di J2) rendono questo aplotipo uno dei marcatori candidati tramite i quali si ipotizza che si possa ricostruire la diffusione della tecnologia agricola durante il Neolitico.
| Popolazione | Sample size | J1 totale | J1 in assenza di P58 | J-P58 (J1c3) | pubblicazione |
| Nord Est Italia | 67 | 0.0% | NA | NA | Battaglia et al. (2008)[38] |
| Italiani | 915 | 0.7% | NA | NA | Capelli et al. (2009)[39] |
| Siciliani | 236 | 3.8% | NA | NA | Di Gaetano et al. (2009)[40] |
G M201
L'aplogruppo G del cromosoma Y si presenta a basse frequenze in più popolazioni, ma risulta distribuito in Europa, Nord Asia e Asia occidentale, Nord Africa, Medio Oriente, India, Sri Lanka e Malesia.
In Europa occidentale e nell'area del Mar Nero l'aplogruppo G si trova in media in circa il 5% della popolazione. La concentrazione scende al di sotto di questa media in Scandinavia, Polonia, Islanda e isole britanniche.
In Tirolo (e Alto Adige) la percentuale G raggiunge l'8% o più; Nelle zone settentrionali ed in Sardegna raggiunge picchi all'11% della popolazione[43]. Analisi genetiche sui resti della mummia del Similaun, meglio nota come Ötzi, hanno dimostrato che apparteneva a una subclade (G-L91) di questo aplogruppo [44].
Romanizzazione
In Europa ed in Italia il fenomeno della romanizzazione ha apportato importanti contributi sul piano culturale, politico e storico delle popolazioni, tuttavia non emerge un ruolo di omogeneizzazione a livello genetico[45]. Le varie aree dell'Italia presentano ad oggi differenze sul piano della distribuzione percentuale degli aplotipi, legati alla discendenza Y nelle varie zone d'Italia, la quale appare più similare alla distribuzione del periodo pre-romano.
Sotto il profilo genetico la penisola italiana e le isole del Mediterraneo hanno interessato diversi autori: ciò si deve alla contemporanea presenza di popoli rimasti relativamente isolati, quali per esempio i Sardi, e popolazioni enormemente eterogenee e da sempre interessate da fenomeni migratori, quali per esempio quelle costiere dell'Italia meridionale[45].

Anche le migrazioni avvenute in suolo italico dalla caduta dell'Impero romano d'Occidente fino all'anno 1000 d.C. non hanno alterato in maniera significativa il pool genetico degli odierni italiani; si stima che l'aplogruppo I sia diffuso fra gli italiani del nord nell'ordine del 2-3% e fra gli italiani del sud nell'1-1,5%[48]. Alcuni subcladi dell'aplogruppo R1a, che potrebbero essere penetrati in Italia assieme alle invasioni provenienti dall'Europa nord-occidentale, compaiono in ordini di percentuali fra il 2,5% ed in particolare R1b1c9 a cui appartiene il 3,5% degli italiani[49], 5,6% nel Nord Italia[12][13], particolarmente diffusa fra i popoli dell'Europa nord-occidentale, essendo però questi subcladi appena citati piuttosto diffusi anche in altre aree europee è difficile stabilire se siano stati veramente introdotti in Italia da migrazioni di quei popoli.
Saraceni
Ulteriori migrazioni di popoli come le scorrerie saracene non hanno intaccato in percentuali significative la distribuzione aplotipica dell'Italia peninsulare, eccezion fatta per la Sicilia dove la civiltà araba e berbera ha prosperato per circa un secolo (emirato di Sicilia) e l'impatto della colonizzazione arabo-berbera è avvenuto in modo più intenso rispetto alla penisola, non modificando tuttavia in modo significativo la composizione neolitica dell'isola[50]. Il contributo berbero si stima tra il 6% e il 7,5% in Sicilia, 6,5% in Puglia Nord-Occidentale, 4,8% in Campania orientale; è da sottolineare il fatto che precedenti studi effettuati nelle stesse aree diedero percentuali minori. Le altre regioni dimostrano percentuali minori, dallo 0 al 2%[51][52].

Albero filogenetico degli aplogruppi del cromosoma Y e X
Mappa Genetica dell'Europa
Recentemente diversi ricercatori hanno contribuito allo sviluppo di una mappa genetica dell'Europa, questa mappa mostra un evidente grado di somiglianza strutturale alla mappa geografica. Le principali differenze genetiche si sono riscontrate fra le popolazioni del nord e del sud.
Una ricerca genetica pubblicata su Current Biology, che prendeva a campione 24 popolazioni umane europee, conferma che le differenze genetiche in Europa sono direttamente proporzionali alla distanza tra i popoli con maggiori differenze per l'Italia e la Finlandia. Le popolazioni abitanti questi due paesi mostrerebbero quindi una maggiore distanza genetica rispetto alle altre 22 popolazioni campionate per via delle barriere geografiche che li separano dalle altre popolazioni.[53][54] Per ciò che concerne l'Italia in particolare uno studio del 2014 rileva che le popolazioni italiane sono estremamente eterogenee da un punto di vista genetico, tanto da poter paragonare la loro diversità a quella che si osserva tra gruppi che vivono agli angoli opposti dell'Europa.[55][56]
I ricercatori ipotizzano tre principali eventi di colonizzazione a partire da sud, avvenuti circa 45.000 anni fa, i primi esseri umani moderni entrerebbero in Europa da sud, dopo questo ingresso si verificò un'interruzione dei flussi migratori dovuto ad un massimo glaciale, circa 20.000 anni fa, la seconda colonizzazione avvenuta al ritiro dei ghiacci risalirebbe a circa 17.000 anni fa a partire da popolazioni di ritorno dalle zone di rifugio a sud[57], l'ultima colonizzazione si ebbe intorno ai 10.000 anni fa con l'espansione dal Vicino Oriente dell'agricoltura.[58][59]
Note
- ^ Ancora oggi alcuni biologi, tra cui Jared Diamond, sono del parere che la distinzione tra i generi Homo e Pan sia del tutto arbitraria e artificiosa, risolvibile riclassificando lo scimpanzé comune come Homo troglodytes ed il bonobo come Homo paniscus.
- ^ L'uomo moderno secondo gli studi genetici, è originario dell'Africa. Durante il processo di migrazione chiamato Out-of-Africa 2.
- ^ SNPs (Single Nucleotide Polymorphisms).
- ^ Non-Recombining Y-chromosome.
- ^ a b Ornella Semino, G.Passarino; PJ. Oefner et al., The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective, in Science Magazine, nº 5494, novembre 2000, pp. 1155-1159.
- ^ [1] Eupedia: Distribution of European Y-chromosome DNA (Y-DNA) haplogroups by region in percentage. See note n° 4.
- ^ [2] American Journal of Human Genetics : Mitochondrial Haplogroup U5b3: A Distant Echo of the Epipaleolithic in Italy and the Legacy of the Early Sardinians
- ^ [3] Distribuzione europea del DNA mitocondriale (mtDNA) aplogruppi per regione in percentuale
- ^ [4] Differenziazione su base cronologica degli aplogruppi del mtDNA
- ^ [5] Siiri Rootsi : Y-Chromosome haplogroup I prehistoric gene flow in Europe
- ^ Di Giacomo et al. Clinal patterns of human Y chromosomal diversity in continental Italy and Greece are dominated by drift and founder effects. Molecular Phylogenetics and Evolution. In Press.
- ^ a b [6] Myres et al. - A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe (2010)
- ^ a b [7] Dienekes blogspot - R1b founder effect in Central and Western Europe (25 agosto 2010)
- ^ Capelli C, Brisighelli F, Scarnicci F, Arredi B, Caglia' A, Vetrugno G, Tofanelli S, Onofri V, Tagliabracci A, Paoli G, Pascali Y chromosome genetic variation in the Italian peninsula is clinal and supports an admixture model for the Mesolithic-Neolithic encounter VL. Mol Phylogenet Evol. 2007 Jul;44(1):228-39. Epub 2006 Dec 13. PMID 17275346
- ^ B. Arredi, E. S. Poloni and C. Tyler-Smith (2007). "The peopling of Europe". In Crawford, Michael H.. Anthropological genetics: theory, methods and applications. Cambridge, UK: Cambridge University Press. p. 394. ISBN 0-521-54697-4.
- ^ Chiaroni, J; Underhill, P; Cavalli-Sforza, L.L. (2009), "Y chromosome diversity, human expansion, drift and cultural evolution", PNAS 106 (48): 20174:20179
- ^ a b Myres, Natalie (2010), "A major Y-chromosome haplogroup R1b Holocene effect in Central and Western Europe", European Journal of Human Genetics, doi:10.1038/ejhg.2010.146
- ^ Cruciani et al. (2010), "Strong intra- and inter-continental differentiation revealed by Y chromosome SNPs M269, U106 and U152", Forensic Science International: Genetics, doi:10.1016/j.fsigen.2010.07.006
- ^ David K. Faux A Genetic Signal of Central European Celtic Ancestry:Preliminary Research Concerning Y-Chromosome Marker U152 17 November 2008 [8]
- ^ David K. Faux Y-Chromosome Marker S28/U152 Haplogroup R-U152
- ^ Mappa di dispersione del sottoclade S28/U152 [9] basata sulla ricostruzione della discendenza Y ISOGG [10]
- ^ Gérard et al.; Berriche, S; Aouizérate, A; Diéterlen, F; Lucotte, G (2006), "North African Berber and Arab influences in the western Mediterranean revealed by Y-chromosome DNA haplotypes", Human Biology 78 (3): 307–316, doi:10.1353/hub.2006.0045, PMID 17216803
- ^ a b Cruciani et al., F.; La Fratta, R.; Trombetta, B.; Santolamazza, P.; Sellitto, D.; Colomb, E. B.; Dugoujon, J.-M.; Crivellaro, F. et al. (2007), "Tracing Past Human Male Movements in Northern/Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12", Molecular Biology and Evolution 24 (6): 1300–1311, doi:10.1093/molbev/msm049, PMID 17351267 Also see Supplementary Data.
- ^ Adams et al.; Bosch, Elena; Balaresque, Patricia L.; Ballereau, StéPhane J.; Lee, Andrew C.; Arroyo, Eduardo; López-Parra, Ana M.; Aler, Mercedes et al. (2008), "The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula", The American Journal of Human Genetics 83 (6): 725, doi:10.1016/j.ajhg.2008.11.007, PMC 2668061, PMID 19061982
- ^ Silva et al.; Carvalho, Elizeu; Costa, Guilherme; Tavares, Lígia; Amorim, António; Gusmão, Leonor (2006), "Y-chromosome genetic variation in Rio De Janeiro population", American Journal of Human Biology 18 (6): 829–837 doi=10.1002/ajhb.20567, doi:10.1002/ajhb.20567, PMID 17039481
- ^ Cruciani et al., Fulvio; La Fratta, Roberta; Santolamazza, Piero; Sellitto, Daniele; Pascone, Roberto; Moral, Pedro; Watson, Elizabeth; Guida, Valentina et al. (May 2004), "Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa" (PDF), American Journal of Human Genetics 74 (5): 1014–1022, doi:10.1086/386294, PMC 1181964, PMID 15042509
- ^ Underhill and Kivisild; Kivisild, Toomas (2007), "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations", Annu. Rev. Genet. 41: 539–64, doi:10.1146/annurev.genet.41.110306.130407, PMID 18076332
- ^ Steven C. Bird Haplogroup E3b1a2 as a Possible Indicator of Settlement in Roman Britain by Soldiers of Balkan Origin http://www.jogg.info/32/bird.htm
- ^ ISOGG
- ^ Ornella Semino et al., "Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area," American Journal of Human Genetics 74:1023–1034, 2004
- ^ R. King and P.A. Underhill (2002), Congruent distribution of Neolithic painted pottery and ceramic figurines with Y-chromosome lineages, Antiquity 76:704-714
- ^ J. Chiaroni et al. (2008), Correlation of annual precipitation with human Y-chromosome diversity and the emergence of Neolithic agricultural and pastoral economies in the Fertile Crescent, Antiquity Volume: 82 Number: 316 Page: 281–289
- ^ F. Di Giacomo et al. (2004), Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe, Human Genetics 115(5):357-71
- ^ Capelli C et. al Y chromosome genetic variation in the Italian peninsula is clinal and supports an admixture model for the Mesolithic-Neolithic encounter Mol Phylogenet Evol. 2007 Jul;44(1):228-39. Epub 2006 Dec 13
- ^ King, R. J.; Ozcan, S. S., Carter, T., Kalfoglu, E., Atasoy, S., Triantaphyllidis, C., Kouvatsi, A., Lin, A. A., Chow, C-E. T., Zhivotovsky, L. A., Michalodimitrakis, M., Underhill, P. A., (2008). "Differential Y-chromosome Anatolian Influences on the Greek and Cretan Neolithic". Annals of Human Genetics 72 Issue 2 March 2008 (Pt 2): 205–214. doi:10.1111/j.1469-1809.2007.00414.x. PMID 18269686
- ^ Y Chromosome Consortium "YCC" (2002), "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups", Genome Research 12 (2): 339–348, doi:10.1101/gr.217602, PMC 155271, PMID 11827954
- ^ Tofanelli, Sergio; Ferri, Gianmarco; Bulayeva, Kazima; Caciagli, Laura; Onofri, Valerio; Taglioli, Luca; Bulayev, Oleg; Boschi, Ilaria et al. (2009), "J1-M267 Y lineage marks climate-driven pre-historical human displacements", European Journal of Human Genetics 17: 1520–1524, doi:10.1038/ejhg.2009.58
- ^ Battaglia; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri et al. (2008), "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe", European Journal of Human Genetics 17 (6): 820–830, doi:10.1038/ejhg.2008.249, PMC 2947100, PMID 19107149
- ^ Capelli; Onofri, Valerio; Brisighelli, Francesca; Boschi, Ilaria; Scarnicci, Francesca; Masullo, Mara; Ferri, Gianmarco; Tofanelli, Sergio et al. (2009), "Moors and Saracens in Europe: estimating the medieval North African male legacy in southern Europe", European Journal of Human Genetics 17 (6): 848–852, doi:10.1038/ejhg.2008.258, PMC 2947089, PMID 19156170
- ^ Di Gaetano; Cerutti; Crobu; Robino (2009), "Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome", European Journal of Human Genetics 17 (1): 91–99, doi:10.1038/ejhg.2008.120, PMC 2985948, PMID 18685561
- ^ Semino O, Passarino G, Oefner PJ, et al. (November 2000). "The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective". Science 290 (5494): 1155–9. doi:10.1126/science.290.5494.1155. PMID 11073453
- ^ Cinnioğlu C, King R, Kivisild T, et al. (January 2004). "Excavating Y-chromosome haplotype strata in Anatolia". Hum. Genet. 114 (2): 127–48. doi:10.1007/s00439-003-1031-4. PMID 14586639
- ^ Zei, G. et al. (2003). "From surnames to the history of Y chromosomes: the Sardinian population as a paradign.". Eur J of Human Genetics 11 (10): 802–07. doi:10.1038/sj.ejhg.5201040. PMID 14512971
- ^ Andreas Keller, Angela Graefen, Markus Ball, Mark Matzas, Valesca Boisguerin, Frank Maixner, Petra Leidinger, Christina Backes, Rabab Khairat, Michael Forster, Björn Stade, Andre Franke, Jens Mayer, Jessica Spangler, Stephen McLaughlin, Minita Shah, Clarence Lee, Timothy T. Harkins, Alexander Sartori, Andres Moreno-Estrada, Brenna Henn, Martin Sikora, Ornella Semino, Jacques Chiaroni, Siiri Rootsi, Natalie M. Myres, Vicente M. Cabrera, Peter A. Underhill, Carlos D. Bustamante e Eduard Egarter Vigl, New insights into the Tyrolean Iceman's origin and phenotype as inferred by whole-genome sequencing, in Nature Communications, vol. 3, 28 febbraio 2012, p. 698, DOI:10.1038/ncomms1701, PMID 22426219.
- ^ a b Cavalli Sforza Luigi L.; Menozzi Paolo; Piazza Alberto, Storia e geografia dei geni umani, Adelphi (collana Gli Adelphi), 2000(2 ed.), EAN 9788845915888.
- ^ [11] Rootsi et al (2004)-Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe (Figure 1 , Map C)
- ^ Balanovsky et al (2008) Figure 4, Map A. Distribution of Y Chromosomal Haplogroups I1a in Europe
- ^ [12] Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe
- ^ [13] Y-chromosome Short Tandem Repeat DYS458.2 Non-consensus Alleles Occur Independently in Both Binary Haplogroups J1-M267 and R1b3-M405
- ^ [14] Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome (2008)
- ^ [15] European Journal of Human Genetics : Moors and Saracens in Europe , estimating the medieval North African male legacy in southern Europe
- ^ [16] Dienekes blogspot : North African male legacy in Southern Europe quantified
- ^ (EN) Nicholas Wade, Visual Science - The Genetic Map of Europe, in N.Y. Times, The New York Times Co., 13 agosto 2008.
- ^ (EN) AA.VV., Correlation between Genetic and Geographic Structure in Europe, in Current Biology, Cell Press, 26 agosto 2008.
- ^ Biodiversità umana, italiani al primo posto
- ^ Capocasa M. et al. Linguistic, geographic and genetic isolation: a collaborative study of Italian populations. Journal of Anthropological Sciences, volume 92, doi 10.4436/JASS.92001
- ^ Con il termine zone di rifugio, si intendono regioni geografiche non meglio identificate nelle quali i primi migranti giunti in europa si sarebbero concentrati in seguito all'avanzamento dei ghiacci, per poi fuoriuscirne ed espandersi al ritiro degli stessi, cioè al termine dell'ultimo massimo glaciale
- ^ Genes mirror geography within Europe
- ^ Genes mirror geography within Europe - Figure 2
![[9]](http://www.eupedia.com/images/content/Haplogroup-R1b-S28.gif){kind=link}
Bibliografia
- Luigi L. Cavalli Sforza, Paolo Menozzi e Alberto Piazza, Storia e geografia dei geni umani, 2ª ed., Milano, Adelphi, 2000, ISBN 978-88-459-1588-8.
- L. e F. Cavalli-Sforza, Chi siamo, Milano, Mondadori, 1993, ISBN 88-04-36901-9.
- Luigi L. Cavalli Sforza e Albert J. Ammerman, La transizione neolitica e la genetica di popolazione in Europa, Torino, Boringhieri, 1986, ISBN 88-339-0043-6.
- Luigi L. Cavalli Sforza, Francesco Cavalli-Sforza e Ada Piazza, Razza o pregiudizio? L'evoluzione dell'uomo fra natura e storia, Milano, Einaudi scuola, 1996, ISBN 88-286-0297-X.
- (EN) Cultural transmission and evolution. A quantitative approach, con Marcus W. Feldman, Princeton, Princeton University Press, 1981.
- (EN) Luigi L. Cavalli-Sforza e A.W.F. Edwards, Analysis of human evolution, in S. J. Geerts (a cura di), Genetics Today. Proceedings of the XI International Congress of Genetics, The Hague, The Netherlands, September, 1963, vol. 3, Oxford, Pergamon Press, 1965, pp. 923–933.
- (EN) The genetics of human population, con Walter F. Bodmer, San Francisco, W. H. Freeman and C., 1971.
Nessun commento:
Posta un commento